
Dans l’École du Prieuré de Arthur Conan Doyle, Sherlock Holmes interroge le Dr. Watson:
– Faites un effort de mémoire, reportez vos souvenirs en arrière. Les voyez-vous, ces empreintes sur le sentier?
– Oui, je les vois.
– Vous vous rappelez qu’elles étaient tantôt ainsi placées, Watson, et il disposa de cette manière des miettes de pain : : : : : : : d’autres fois ainsi. : . : . : . : d’autres comme ceci . . . . . . . . , vous rappelez-vous tout cela?
– Non, cela m’est impossible.
– Moi, c’est différent, j’en jurerais, mais nous irons les revoir. Que j’ai donc été aveugle de ne pas en tirer plus tôt une conclusion !
– Laquelle?
– Seulement celle-ci: c’est une vache remarquable que celle qui peut ainsi aller au pas, au trot et au galop. Allons donc un peu à la découverte!
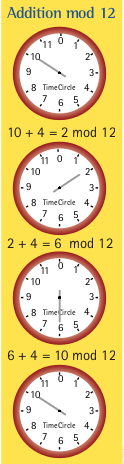 Comme Holmes et Watson, allons à la découverte des allures animales, c’est-à-dire, des différents modes de déplacement. Nous allons concentrer nos efforts sur les symétries des allures et notre outil principal sera la théorie des groupes.
Comme Holmes et Watson, allons à la découverte des allures animales, c’est-à-dire, des différents modes de déplacement. Nous allons concentrer nos efforts sur les symétries des allures et notre outil principal sera la théorie des groupes.
Une allure est un mouvement périodique dans le temps. Par exemple, chez l’humain, la marche consiste en une alternance gauche-droite dans le mouvement des jambes, répété de façon identique à chaque pas. Supposons qu’un pas complet, allant du contact initial du pied gauche au sol au contact suivant du pied gauche avec le sol, dure une seconde, ceci détermine la période de l’allure. Donc, par une translation temporelle par un multiple d’une seconde, nous pouvons ramener tous les pas complets, vers le premier pas initial et étudier une seule période que nous nommerons un cycle et les temps à l’intérieur d’un cycle sont appelés les phases.
Si \(\phi\) est une phase d’un cycle d’une seconde, alors \(\phi\) est un élément de l’intervalle semi-ouvert [0, 1[. Nous pouvons additionner les phases modulo 1. Puisque cette addition est différente de l’addition normale, nous la désignons avec son propre symbole ⊕ et l’addition modulo 1 de phases s’écrit
\[\phi_1 \oplus \phi_2:= (\phi_1 + \phi_2) \mod 1.\]
 L’ensemble [0, 1[ avec l’opération ⊕ forme un groupe que l’on note par S1. C’est le groupe de rotations sur un cercle de longueur 1 de telle façon que si la somme dépasse 1, alors on ne garde que le reste de la division par 1. C’est le genre d’addition que nous faisons avec une horloge de 12 heures1, par exemple si on ajoute 6 heures à 8h00, on dit qu’il est 2h00. En effet 8 + 6 = 14 et la division par 12 nous donne 2. Cette opération est appelée addition modulo n, on la désigne en ajoutant le symbole mod n, où n est le nombre naturel de la division. Ci-haut, nous avons mod 1 et mod 12. Pour le vérifier il suffit de montrer que toutes les propriétés d’un groupe sont satisfaites. Dans l’exemple de la marche, nous avons pris le contact initial du pied gauche au sol comme début du cycle, mais ce choix est arbitraire et tout autre moment du cycle est tout aussi approprié comme moment initial. Nous avons donc une symétrie sur les phases donné par l’ensemble des translations de phases, c’est-à-dire S1 = ([0, 1[; ⊕) est le groupe de symétries d’un cycle.
L’ensemble [0, 1[ avec l’opération ⊕ forme un groupe que l’on note par S1. C’est le groupe de rotations sur un cercle de longueur 1 de telle façon que si la somme dépasse 1, alors on ne garde que le reste de la division par 1. C’est le genre d’addition que nous faisons avec une horloge de 12 heures1, par exemple si on ajoute 6 heures à 8h00, on dit qu’il est 2h00. En effet 8 + 6 = 14 et la division par 12 nous donne 2. Cette opération est appelée addition modulo n, on la désigne en ajoutant le symbole mod n, où n est le nombre naturel de la division. Ci-haut, nous avons mod 1 et mod 12. Pour le vérifier il suffit de montrer que toutes les propriétés d’un groupe sont satisfaites. Dans l’exemple de la marche, nous avons pris le contact initial du pied gauche au sol comme début du cycle, mais ce choix est arbitraire et tout autre moment du cycle est tout aussi approprié comme moment initial. Nous avons donc une symétrie sur les phases donné par l’ensemble des translations de phases, c’est-à-dire S1 = ([0, 1[; ⊕) est le groupe de symétries d’un cycle.
Groupe symétrique
 Les permutations des éléments d’un ensemble forment, avec l’opération de composition, ce qu’on appelle un groupe symétrique. Pour un ensemble de n éléments, le groupe symétrique est désigné par Sn et il contient n! éléments. L’illustration ci-contre représente huit des vingt-quatre éléments du groupe des permutations d’un ensemble de quatre éléments. Par exemple, la permutation \(\sigma_2\) est telle que
Les permutations des éléments d’un ensemble forment, avec l’opération de composition, ce qu’on appelle un groupe symétrique. Pour un ensemble de n éléments, le groupe symétrique est désigné par Sn et il contient n! éléments. L’illustration ci-contre représente huit des vingt-quatre éléments du groupe des permutations d’un ensemble de quatre éléments. Par exemple, la permutation \(\sigma_2\) est telle que
1→3, 2→4 et 3→1 et 4 → 2.
Avant de voir comment s’effectue la composition, on peut alléger la représentation graphique et la notation, comme l’illustre la deuxième figure à gauche. Si on considère la permutation de 4 éléments donnée par \(\sigma_a\), on voit que les éléments 1, 2 et 3 sont permutés entre eux de façon circulaire et que l’élément 4 n’est pas déplacé. On adopte la notation plus compacte \(\sigma_a\) = (1 2 3) où (1 2 3) indique que 1→2 et 2→3 et 3→1 et l’absence de 4 indique qu’il n’est pas déplacé, 4→4.
 Composition de permutations
Composition de permutations
Voyons maintenant comment se fait la composition en considérant, dans le groupe S4, la composition (1324) * (1324): commençant par le terme de droite nous avons 1→3, ensuite par le terme de gauche nous avons 3→2. Au total, ceci implique 1→2. Nous recommençons, par le terme de droite 2→4, suivi par le terme de gauche indiquant 4→1, au total 2→1. Le même raisonnement implique 3→4 et 4→3. Donc,
\[\sigma^2 =\sigma*\sigma=(1\,2)(3\,4),\]
\(\sigma^3 = (1\,4\,2\,3)\) et \(\sigma^4\) est l’identité (1 2 3 4),désignée par e.
 Générateurs d’un groupe
Générateurs d’un groupe
Un groupe (G; *) peut être décrit par ses générateurs qui sont les éléments qui engendrent tous les autres éléments de G par l’opération *. Par exemple, en prenant la permutation circulaire \(\sigma\)=(1 2 3 4), la composition de \(\sigma\) avec elle-même engendre un groupe de quatre éléments, noté \(\mathbb{Z}_4.\)
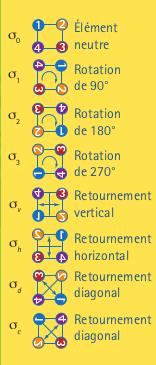 Le groupe S4, quant à lui,est engendré par les permutations (1 2), (2 3) et (3 4). Le groupe de 8 éléments engendré par les deux permutations (1 2 3 4) et (1 4) (2 3) se nomme le groupe dihédral et a comme symbole: D4. En plus des quatre éléments engendrés par (1 2 3 4), ce groupe a comme éléments: (1 4) (2 3), (2 4), (1 2) (3 4) et (1 3). Dans le tableau à droite, on a représenté les huit symétries du tableau en haut à gauche. Ce sont les huit symétries du groupe D4. Les quatre premières sont celles du groupe \(\mathbb{Z}_4.\)
Le groupe S4, quant à lui,est engendré par les permutations (1 2), (2 3) et (3 4). Le groupe de 8 éléments engendré par les deux permutations (1 2 3 4) et (1 4) (2 3) se nomme le groupe dihédral et a comme symbole: D4. En plus des quatre éléments engendrés par (1 2 3 4), ce groupe a comme éléments: (1 4) (2 3), (2 4), (1 2) (3 4) et (1 3). Dans le tableau à droite, on a représenté les huit symétries du tableau en haut à gauche. Ce sont les huit symétries du groupe D4. Les quatre premières sont celles du groupe \(\mathbb{Z}_4.\)
La notion générale de groupe
Un groupe est une structure algébrique qui consiste en une paire $(G; ∗)$ où $G$ est un ensemble, ∗ est une opération et telle que $G$ est fermé sous l’opération, c’est-à-dire
\[a ∗ b \in G \: \text{pour tout} \: a, b \in G.\]
De plus les trois propriétés suivantes sont satisfaites:
1. associativité:
\[a ∗ (b ∗ c) = (a ∗ b) ∗ c\]
2. élément neutre: il existe $e \in G$ tel que:
\[e ∗ a = a ∗ e = a \: \text{pour tout} \: a \in G\]
3. inverse: pour tout $a \in G,$ il existe un unique élément $b$ tel que
\[a ∗ b = b ∗ a = e.\]
Les allures des bipèdes
 Nous sommes maintenant prêts à formuler une classification des allures de bipèdes selon leurs groupes de symétries. Pour simplifier la classification, nous allons seulement tenir compte du patron de contacts des pieds au sol. Nous supposons un cycle d’une seconde et comme phase initiale nous choisirons le moment de contact du pied gauche. Commençons par le bond qui correspond à une impulsion simultanée des deux jambes, c’est l’allure naturelle des kangourous. Les pieds gauche et droit touchent le sol simultanément, et donc la différence de phase gauche-droit dans le patron des contacts au sol est de zéro; ceci est illustré à l’aide d’un diagramme comme à la figure suivante. La marche et la course chez le bipède ont un diagramme de contacts au sol données dans la figure ci-contre.
Nous sommes maintenant prêts à formuler une classification des allures de bipèdes selon leurs groupes de symétries. Pour simplifier la classification, nous allons seulement tenir compte du patron de contacts des pieds au sol. Nous supposons un cycle d’une seconde et comme phase initiale nous choisirons le moment de contact du pied gauche. Commençons par le bond qui correspond à une impulsion simultanée des deux jambes, c’est l’allure naturelle des kangourous. Les pieds gauche et droit touchent le sol simultanément, et donc la différence de phase gauche-droit dans le patron des contacts au sol est de zéro; ceci est illustré à l’aide d’un diagramme comme à la figure suivante. La marche et la course chez le bipède ont un diagramme de contacts au sol données dans la figure ci-contre.
Identifions les contacts au sol: 1-gauche et 2-droite. Le bond a une symétrie spatiale évidente qui est la permutation gauche-droite, donc \(\kappa = (1 2)\) et son groupe de symétrie est noté \(\mathbb{Z}_2(\kappa):= \{1, \kappa \}.\) La marche et la course ont plutôt une symétrie spatio-temporelle, c’est-à-dire que l’action d’une permutation est équivalente à un déplacement de phase. La permutation (1 2) correspond à avancer par une phase de 0,5 dans le cycle. On note par \((\kappa, 1/2)\) la combinaison de la symétrie spatiale et de la translation de phase par 0,5. Le groupe de symétrie de la marche (et de la course) est donc
\[\mathbb{Z}_2 = \displaystyle \left \{ 1, (\kappa, 1/2) \right \}.\]
 Les allures des quadrupèdes
Les allures des quadrupèdes
Le doublement du nombre de pattes par rapport aux bipèdes amène une bien plus grande variété d’allures. Nous connaissons bien quelques allures de quadrupèdes; par exemple, le trot et le galop. Or, à l’oeil nu, il est pratiquement impossible de suivre l’évolution du mouvement des jambes et d’autant plus, de dé- terminer les phases de contact des pieds au sol. Edweard Muybridge (1830-1904) est l’inventeur de la photographie à haute vitesse et fut le premier à cataloguer les allures de quadrupèdes grâce à ses techniques. Il est un précurseur de la cinématographie. Par la suite, plusieurs zoologistes tels Hildebrand2 et Gambaryan3 ont étudié les phases de contact au sol chez plusieurs espèces de quadrupèdes et ont identifié (partiellement) leurs symétries spatiales.
En 1872, Leland Stanford alors gouverneur de la Californie et amateur de courses de chevaux était d’avis que lors du trot et du galop d’un cheval, les quatre pattes ne touchaient pas au sol pendant une partie du cycle. Il engagea Muybridge pour faire une étude photo- graphique de ce problème, ce qui amena celui-ci à imaginer un dispositif par lequel une série d’appareils photos placés à intervalles réguliers se déclenchaient au passage du cheval. Il put de cette façon confirmer la thèse de Stanford.
Muybridge perfectionna ses techniques et produisit un catalogue impressionnant d’allures de quadrupèdes. La figure à gauche montre les allures de quadrupèdes les plus fréquemment observées ainsi que les différences de phase du contact des pieds au sol, avec le pied postérieur gauche fixé à la phase 0.
La marche (le « pas » que mentionne Holmes) est utilisé par la plupart des quadrupèdes à basse vitesse, le bond est utilisé par les écureuils et autres rongeurs, le pas est une allure utilisée par les chameaux. Les galops sont utilisés par la plupart des espèces pour atteindre des vitesses supérieures.
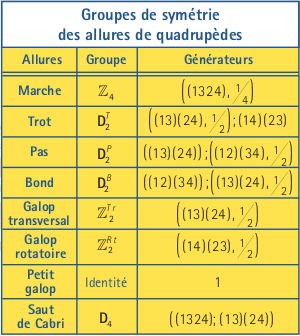
Pour simplifier notre tâche, nous allons nous limiter aux permutations qui se trouvent dans le groupe dihédral D4. Le tableau de la page suivante présente les symétries spatio-temporelles des allures à la figure à gauche. Dans le tableau, les générateurs (1324) et (1 3)(2 4) engendrent un groupe similaire en structure au groupe D4 ci-haut, mais avec une représentation graphique différente.
Commençons par le saut de cabri. Toutes permutations des jambes laissent les phases inchangées, donc D4 est le groupe de symétries du saut de cabri. Le groupe de symétries du trot est engendré par les symétries suivantes: la permutation gauche-droite (13)(24) avec une translation de phase 1/2 et la permutation des jambes sur une même diagonale (14)(23). Le saut de cabri est observé particulièrement chez les antilopes et autres ongulés, surtout pour échapper rapidement à leurs prédateurs. C’est aussi l’allure préférée du fameux « Pepé le Pew » des Looney Tunes (mais dans ce cas-ci, c’est lui le prédateur…).
 Brisures de symétries
Brisures de symétries
Nous voyons dans les allures de quadrupèdes que certains groupes de symétrie sont inclus dans certains autres; c’est-à-dire qu’ils sont sous-groupes d’autres groupes. La figure à droite montre un diagramme des groupes de symétries de quadrupèdes avec les inclusions indiquées par les flèches. Les différentes allures sont généralement observées pour des intervalles de vitesses ne s’intersectant que peu. Par exemple, beaucoup de quadrupèdes préfèrent la marche à basse vitesse, pour ensuite changer à un trot à une vitesse intermédiaire et finalement, brisent le trot pour un galop (soit rotatoire ou transversal) aux vitesses maximales. En observant le diagramme de sous-groupes, la transition trot-galop s’accompagne donc d’une brisure de symétrie. Si on pousse l’idée de brisure de symétrie à l’extrême, la locomotion elle-même peut-être vue comme une brisure de symétrie. En effet, considérons un animal immobile debout, c’est un état d’équilibre, c’est-à-dire qu’il est invariant par toutes translations dans le temps, il est le même au temps t qu’au temps t + s pour tout \(s \in \mathbb{R}.\) Le groupe de symétrie de cet état est \((\mathbb{R}; +)\) où le + est l’addition ordinaire. En amorçant une locomotion (disons de période 1) la symétrie \((\mathbb{R}; +)\) est donc brisée pour la symétrie d’un mouvement périodique S1 = ([0, 1[; ⊕).
Symétries et neurones
 La classification des allures par groupes de symétries n’est pas seulement élégante, mais elle permet aussi d’émettre des hypothèses sur l’architecture de réseaux de neurones produisant la locomotion. Le Centre de Programmes Moteurs (CPM) de locomotion est un réseau de neurones du système nerveux central qui génère et coordonne les influx électriques rythmiques vers les muscles des jambes, produisant ainsi les allures. L’identification des neurones d’un CPM chez les vertébrés, et en particulier les mammifères, est un problème complexe de physiologie animale. Même lorsque certains neurones sont connus, comprendre la dynamique du CPM est un formidable défi. Il est donc naturel d’explorer la question de l’architecture d’un CPM par la modélisation. Ceci a mené à des découvertes surprenantes sur la possible structure du CPM non seulement pour les quadrupèdes, mais aussi à tous les multipèdes. En effet, le modèle minimal de neurones couplés avec une architecture symétrique pouvant produire les allures périodiques de la marche, du trot et du pas distinctement du point de vue des symétries et de manière robuste doit nécessairement avoir huit unités neuronales, c’est-à-dire le double du nombre de pattes. Une interprétation plausible est qu’une moitié du réseau contrôle les muscles flexeurs et l’autre moitié les muscles extenseurs. Les détails de ce chapitre de l’histoire requièrent des notions de théorie des groupes et des systèmes dynamiques pas du tout élémentaire, même pour monsieur Holmes4.
La classification des allures par groupes de symétries n’est pas seulement élégante, mais elle permet aussi d’émettre des hypothèses sur l’architecture de réseaux de neurones produisant la locomotion. Le Centre de Programmes Moteurs (CPM) de locomotion est un réseau de neurones du système nerveux central qui génère et coordonne les influx électriques rythmiques vers les muscles des jambes, produisant ainsi les allures. L’identification des neurones d’un CPM chez les vertébrés, et en particulier les mammifères, est un problème complexe de physiologie animale. Même lorsque certains neurones sont connus, comprendre la dynamique du CPM est un formidable défi. Il est donc naturel d’explorer la question de l’architecture d’un CPM par la modélisation. Ceci a mené à des découvertes surprenantes sur la possible structure du CPM non seulement pour les quadrupèdes, mais aussi à tous les multipèdes. En effet, le modèle minimal de neurones couplés avec une architecture symétrique pouvant produire les allures périodiques de la marche, du trot et du pas distinctement du point de vue des symétries et de manière robuste doit nécessairement avoir huit unités neuronales, c’est-à-dire le double du nombre de pattes. Une interprétation plausible est qu’une moitié du réseau contrôle les muscles flexeurs et l’autre moitié les muscles extenseurs. Les détails de ce chapitre de l’histoire requièrent des notions de théorie des groupes et des systèmes dynamiques pas du tout élémentaire, même pour monsieur Holmes4.
- Voir aussi dans Accromath vol, 7, hiver-printemps, p. 30 et 31 ↩
- M. Hildebrand, Symmetrical gaits of horses, Science, 150, 701-708 (1965). ↩
- P.P. Gambaryan, How Mammals Run: Anatomical Adaptations, Wiley, New-York (1974). ↩
- I. Stewart, The Mathematics of Life, Basic Books.
M. Golubitsky et al. Symmetry in locomotor central pattern generators and animal gaits. Nature 401, 693 – 695 (1999). ↩