
En nous penchant sur les motifs des coquillages, nous allons découvrir qu’il existe un lien avec les motifs des pelages des animaux, la disposition des feuilles sur la tige d’une plante et l’apparition des membres ou organes dans le développement d’un embryon.
En fait, les scientifiques ne comprennent pas bien les mécanismes derrière ces phénomènes. Ils proposent donc des modèles et comparent les résultats des simulations des modèles avec les observations expérimentales.  Lorsque les simulations d’un modèle ressemblent beaucoup aux observations expérimentales, ils concluent que le modèle est une explication plausible du phénomène. Le fait remarquable avec ces différents phénomènes est qu’un même type de modèle permet de fournir simultanément des explications plausibles pour plusieurs d’entre eux. Ce modèle, d’origine chimique, a été proposé par Alan Turing en 1952 dans un article célèbre, « The chemical basis of morphogenesis ».
Lorsque les simulations d’un modèle ressemblent beaucoup aux observations expérimentales, ils concluent que le modèle est une explication plausible du phénomène. Le fait remarquable avec ces différents phénomènes est qu’un même type de modèle permet de fournir simultanément des explications plausibles pour plusieurs d’entre eux. Ce modèle, d’origine chimique, a été proposé par Alan Turing en 1952 dans un article célèbre, « The chemical basis of morphogenesis ».  Il fait intervenir deux substances chimiques ou morphogènes, l’une appelée activateur, et l’autre inhibiteur, qui réagissent l’une avec l’autre, et qui diffusent au travers des cellules. On parle donc de réaction-diffusion. Nous reviendrons sur ce type de modèle, mais nous allons commencer par scruter les motifs du coquillage bien particulier, Cymbiola innexa REEVE, et essayer d’imaginer la démarche d’un scientifique. Ce coquillage remarquable a des motifs très complexes qui vous rappelleront peut-être ceux du triangle de Sierpinski1.
Il fait intervenir deux substances chimiques ou morphogènes, l’une appelée activateur, et l’autre inhibiteur, qui réagissent l’une avec l’autre, et qui diffusent au travers des cellules. On parle donc de réaction-diffusion. Nous reviendrons sur ce type de modèle, mais nous allons commencer par scruter les motifs du coquillage bien particulier, Cymbiola innexa REEVE, et essayer d’imaginer la démarche d’un scientifique. Ce coquillage remarquable a des motifs très complexes qui vous rappelleront peut-être ceux du triangle de Sierpinski1.
Intriguant n’est-ce pas ?
 Nous allons voir comment construire un modèle qui nous génère le triangle de Sierpinski, et comment l’adapter pour produire des motifs comme celui en haut de la page qui ressemble étrangement au motif de Cymbiola innexa REEVE.
Nous allons voir comment construire un modèle qui nous génère le triangle de Sierpinski, et comment l’adapter pour produire des motifs comme celui en haut de la page qui ressemble étrangement au motif de Cymbiola innexa REEVE.
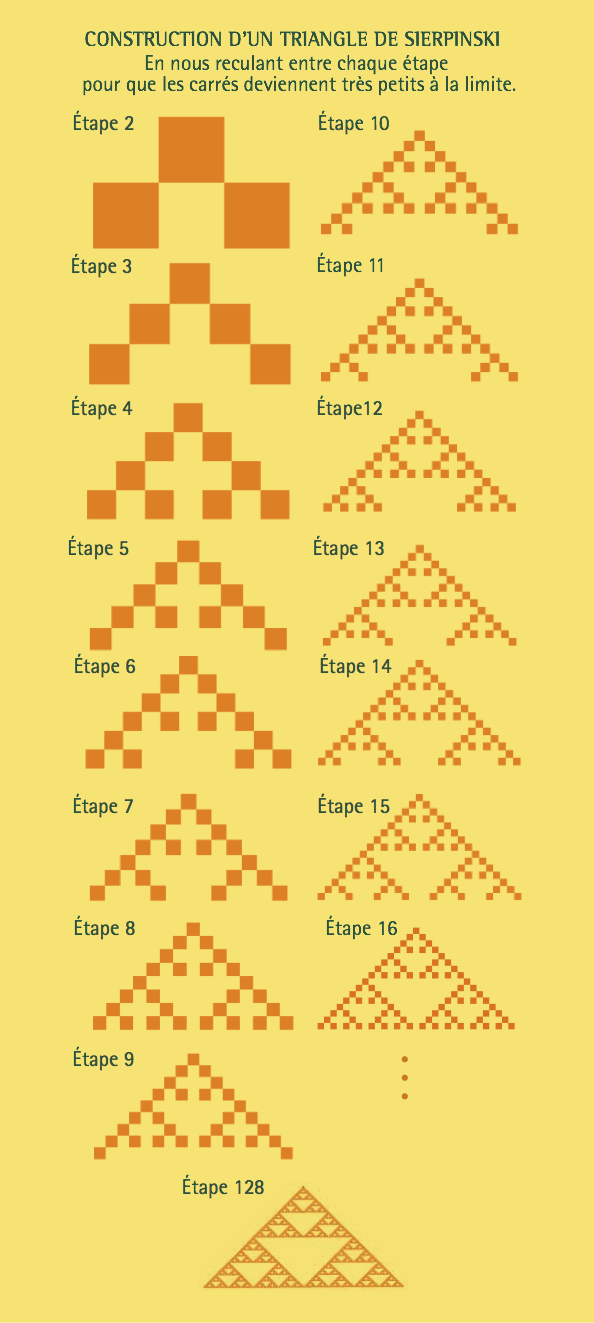
Voyons d’abord comment générer un triangle de Sierpinski infiniment long2. On commence par se donner les règles suivantes de l’addition modulo 2: $$0 + 0 = 0$$ $$0 + 1 = 1 + 0 = 1$$ $$1 + 1 = 0.$$ On va considérer un pavage du plan par des petits carrés de même côté. À chaque carré on attribue, soit la valeur 0, soit la valeur 1. Un carré sera orange si on lui attribue la valeur 1, et jaune sinon. Dans la rangée supérieure on met un carré orange et tous les autres carrés sont jaunes. À partir de cela, on va déterminer une règle de coloriage pour le demi-plan situé sous cette rangée. La règle est la suivante. La couleur de chaque carré est la somme modulo 2 des valeurs numériques attribuées aux couleurs des deux carrés de la rangée supérieure qui le touchent par le coin. L’illustration ci-contre donne les images à diverses étapes de la procédure jusqu’à une image avec 128 rangées, qui ressemble à un triangle de Sierpinski infini vers le bas.
Un système de réaction-diffusion
Commençons par expliquer le principe de la diffusion de l’activateur. Ce sera la même chose pour l’inhibiteur. La densité de l’activateur dans un fonction $a(x, t)$ qui dépend de la position $x$ et du temps $t$. Rappelons que le motif se crée, une strate linéaire à la fois. On appelle $x$ la coordonnée sur cette strate que l’on dessinera horizontale. La coordonnée t désigne le temps. Lorsque t varie on change de strate en bougeant verticalement. On veut connaître comment varie $a(x, t)$ en fonction du temps. Le taux de variation est donné par la dérivée partielle $\frac{\partial a}{\partial t}$.
Sous quelle condition ce taux de variation sera-t-il non nul ? Si la variation $\frac{\partial a}{\partial x}$ est constante, alors $a$ dépend de $x$ et on a autant d’activateur qui pénètre par un côté qu’il en sort par l’autre; d’où la densité ne changera pas en fonction du temps. Donc, pour que $\frac{\partial a}{\partial t}$ soit non nul, il faut que $\frac{\partial a}{\partial x}$ ne soit pas constante et donc, que $\frac{\partial^{2} a}{\partial x^{2}}$ ne soit pas nul : une équation de diffusion a donc toujours un terme en $\frac{\partial^{2} a}{\partial x^{2}}$. L’autre facteur qui fait varier $a$ est l’interaction avec l’inhibiteur que nous appellerons $i$. On obtient donc pour $a$ une équation aux dérivées partielles de la forme $$\frac{\partial a}{\partial t} = A \frac{\partial^{2} a}{\partial x^{2}} + f(a, i)$$ et une équation du même type pour $i$ : $$\frac{\partial i}{\partial t} = B \frac{\partial^{2} i}{\partial x^{2}} + g(a, i)$$
Hans Meinhardt3 a étudié des modèles de ce type pour différentes valeurs des paramètres $A$ et $B$, et différents types de fonctions $f(a, i)$ et $g(a, i)$ et il a pu obtenir des motifs qui ressemblaient beaucoup aux motifs de plusieurs coquillages remarquables, dont Cymbiola Innexa REEVE. Pour des motifs aussi complexes que Cymbiola innexa REEVE il a dû prendre des fonctions $f (a, i)$ et $g(a, i)$ qui dépendaient aussi périodiquement du temps, l’oscillation périodique venant d’un autre couple de substances en réaction-diffusion. Mais, on ne voit toujours pas le lien avec la formule de récurrence ! Ce lien apparaît si on se demande comment on calcule une dérivée seconde. Pour la dérivée première on a approximativement $$\frac{\partial a}{\partial x} \sim \frac{1}{\Delta x}(a (x+\frac{\Delta x}{2}, t) – a(x – \frac{\Delta x}{2}, t)).$$ Par analogie, on aura $$\frac{\partial^{2} a}{\partial x^{2}} \sim \frac{1}{\Delta x^{2}}(a(x + \Delta x,t) – 2a(x,t) + a(x-\Delta x, t)).$$ Si l’on travaille modulo $2$ à cause de la règle $1 + 1 = 0$, et que l’on sait que la dérivée seconde $\frac{\partial^{2} a}{\partial x^{2}}$ par rapport à $x$ est reliée à $\frac{\partial a}{\partial t}$, alors on commence par obtenir quelque chose qui ressemble à la formule de récurrence. L’explication vous paraitra peut-être un peu boiteuse mais, tant la formule de récurrence que la simulation du modèle d’équations de réaction-diffusion de Meinhardt donnent des motifs qui ressemblent suffisamment à ceux de Cymbiola innexa REEVE pour voir dans le modèle proposé un pouvoir non seulement descriptif, mais aussi explicatif du motif observé.
Léopard ou du cou de la girafe, le motif qui se développe dépend de la taille et de la forme de la surface à ce moment-là. Ainsi, on peut avoir une queue rayée avec un corps tacheté, mais pas le contraire. Le tigre et le léopard ont des motifs différents parce que la pigmentation se produit, pour l’un, quand l’embryon est petit, et pour l’autre, quand il est grand (voir texte de Stéphane Durand4).
Des pelages et coquillages aux embryons et aux feuilles des plantes

Oliva porphyria
Lors des premières divisions cellulaires, toutes les cellules de l’embryon sont identiques, et l’embryon, de forme sphérique. Puis, on voit apparaître des amorces de membres, organes, etc., et les cellules se différencient. L’hypothèse sur laquelle les biologistes travaillent remonte à l’article d’Alan Turing (voir page suivante), sur l’origine chimique de la morphogénèse, soit l’ensemble des lois qui déterminent le développement de la forme et la structure des tissus, des organes et des organismes. Turing a émis l’hypothèse qu’au départ deux réactants chimiques sont en équilibre stable et qu’il n’y a pas de motif. Mais les réactants se mettent à diffuser. Cette diffusion rend l’équilibre instable. En fait, un rien déstabilise le système qui perd sa symétrie. L’analogie couramment donnée est celle avec les dunes de sable. Pourquoi se forment-elles sous l’action du vent ? Prenons au départ une surface de sable presque parfaitement plane et faisons souffler le vent à sa surface. Il suffit de quelques petites irrégularités pour que celles-ci arrêtent d’autres grains de sable. Les irrégularités grossissent, et plus elles sont grosses, plus elles ralentissent le sable, ce qui les fait encore grossir. C’est à cause de cette instabilité qu’on n’observe jamais dans la nature des paysages de sable plats, mais plutôt des dunes. Cette même explication de Turing est utilisée pour la formation des motifs, puisque la position d’équilibre pour laquelle les réactants sont uniformément distribués est instable. C’est aussi l’explication sur laquelle travaillent les scientifiques pour les différentes phases de l’embryogénèse, dont la gastrulation et l’organogénèse, lorsque les cellules ou les régions de l’embryon se différencient. Ces différenciations se produiraient au moment où la diffusion déstabilise l’équilibre des réactants chimiques. Il en est de même pour la phyllotaxie, soit l’ordre des feuilles sur la tige d’une plante, ou encore la disposition des éléments d’un fruit ou d’une fleur. La naissance d’une feuille pourrait être expliquée par la déstabilisation de la symétrie cylindrique de la tige dans un processus de réaction-diffusion. La validation biologique reste encore à faire, mais le modèle est très séduisant.
Pour terminer, mentionnons que Hans Meinhardt a proposé des modèles de réaction-diffusion expliquant les motifs de beaucoup d’autres coquillages, dont Conus marmoreus, Oliva porphyria, ou encore Lioconcha castrensis.
Alan Mathison Turing (1912-1954)
 Alan Mathison Turing était un mathématicien anglais, né à Londres le 23 juin 1912. Son père était un fonctionnaire d’administration coloniale. Un an après la naissance, sa mère alla rejoindre son père aux Indes, laissant le jeune Alan à des amis qui se chargèrent de son éducation.
Alan Mathison Turing était un mathématicien anglais, né à Londres le 23 juin 1912. Son père était un fonctionnaire d’administration coloniale. Un an après la naissance, sa mère alla rejoindre son père aux Indes, laissant le jeune Alan à des amis qui se chargèrent de son éducation.
Dès ses débuts à l’école, il se démarque par son intérêt pour les sciences et les mathématiques. Sa carrière de mathématicien débuta au King’s College de l’Université de Cambridge en 1931. En recevant son diplôme , il fut fait « fellow » du King’s College. Durant les années 1937-1938, il s’installe à l’Université Princeton où il travaille sous la direction d’Alonzo Church. Il y développe ce que Church a appelé la « machine de Turing ».
Cette invention s’inscrit dans le contexte du débat sur la notion de « calculabilité »: qu’est-ce qu’une fonction calculable ? C’est une fonction pour laquelle on peut donner un algorithme qui la calcule. Mais, qu’est-ce qu’un algorithme ? C’est un ensemble de règles de calcul qu’on peut appliquer mécaniquement. Si on peut les appliquer mécaniquement, on peut imaginer construire une machine qui va les exécuter. À l’époque, Turing avait déjà développé des machines pour résoudre des problèmes spécifiques : à chaque problème sa machine. Quant à elle, la machine de Turing, une machine théorique, est universelle. À la manière des ordinateurs modernes qu’elle a permis d’entrevoir, elle peut effectuer n’importe quel algorithme sur n’importe quelle entrée : il suffit de coder les instructions de l’algorithme, ainsi que l’entrée, dans un format adéquat.
Ce concept était révolutionnaire pour l’époque. Ce qui est d’autant plus remarquable, c’est que la machine de Turing, si puissante sur le plan théorique, est très simple. Elle consiste en un ruban infini qui est une suite de cases dans lesquelles on inscrit un nombre fini de symboles dans un nombre fini de cases. Chaque programme consiste en un nombre fini d’instructions. Au départ, l’entrée est inscrite dans les cases du ruban. La machine est munie d’un pointeur qui regarde une case à la fois. Sur cette case, il effectue une instruction du programme si une instruction pertinente existe, puis passe à une case adjacente. Il s’arrête quand il ne peut plus effectuer d’instruction, et on lit le résultat du calcul sur le ruban.
Durant la deuxième guerre mondiale, Alan Turing utilisa ses compétences mathématiques dans le décodage des messages allemands, encodés par la machine Enigma qui modifiait continuellement la clé d’encodage des messages. Il fut un des acteurs principaux de ces travaux menés à Bletchley Park, le centre secret du service britannique du chiffre. En 1942, il retourne aux États-Unis pour tenter de percer le secret des codes japonais. Il y rencontre Claude Shannon qui développait la théorie de l’information.
En 1952, Turing a élaboré un modèle biomathématique de la morphogenèse, tant chez l’animal que chez le végétal. Il fit paraître un article, « The chemical basis of morphogenesis » (Philosophical Transactions of the Royal Society of London, août 1952), où il propose trois modèles de formes (Turing patterns). Dans les années 1990, des expériences de chimie ont confirmé expérimentalement les modèles théoriques de Turing.
Après avoir reconnu son homosexualité, en 1952, il fut l’objet de persécutions et sa carrière fut brisée. Il fut écarté des plus grands projets scientifiques, même si ses compétences avaient été reconnues en 1951 alors qu’il est devenu membre de la Royal Society.
Il fut accusé d’indécence et de perversion sexuelle et subit un procès très médiatisé, et comme on hésitait à le condamner à la prison à cause des services rendus durant la guerre, on lui offrit le choix entre l’incarcération et une castration chimique, réduisant sa libido. Il choisit la castration. En 1954, il meurt d’un empoisonnement au cyanure. L’enquête qui s’ensuivit conclut au suicide. Selon certaines sources, le moyen d’ingestion du poison aurait été une pomme partiellement mangée retrouvée près du corps de Turing et qui aurait été imbibée de cyanure, mais on n’a aucune certitude car la pomme n’a pas été analysée. Andrew Hodges qui a écrit la biographie de Turing a émis l’hypothèse que celui-ci a choisi ce mode d’ingestion précisément pour laisser à sa mère la possibilité de croire à un accident. Nombreux sont ceux qui ont souligné le lien entre sa méthode de suicide et le film Blanche-Neige et les Sept Nains, duquel il avait particulièrement apprécié la scène où la sorcière crée la pomme empoisonnée, au point de régulièrement chantonner les vers prononcés par la sorcière: « Plongeons la pomme dans le chaudron, pour qu’elle s’imprègne de poison ».
- Voir l’article « Point fixe de Banach » dans Accromath, hiver-printemps 2010, page 20. ↩
- Nous avons trouvé ce modèle dans un document non signé de l’université de Provence intitulé « Les formes fractales dans la nature ». ↩
- H. Meinhardt, The algorithmic beauty of sea shells, Springer, 1995 ↩
- http://www.crm.umontreal.ca/math2000-1/pub/leopard.html ↩